 |
Artikel bereits veröffentlicht in: Zeitschrift für Feldherpetologie 18: 181–198, Oktober 2011
В© Laurenti-Verlag, Bielefeld, www.laurenti.de
Verbreitung und genetische Herkunft verschiedener Formen der Mauereidechse (Podarcis muralis) in Stuttgart
GUNTRAM DEICHSEL, AXEL KWET & ALBIA CONSUL
Distribution and genetic origins of different forms of the common wall lizard (Podarcis muralis) within the city of Stuttgart, Germany
In Stuttgart there is a strikingly broad phenotypic heterogeneity of common wall lizards, giving rise to the suggestion that the various locations are occupied by alien common wall lizards of different origins. Hence we investigated important sites where such lizards occur in the period between 2007–2010. The purpose of our investigations was to gather data on the genetic origins of Stuttgart’s common wall lizards. We took tissue samples from twenty animals and determined their haplotypes by means of mtDNA analysis. We found that western city quarters are inhabited by P. m. maculiventris (Western or synonymously Southern Alps lineage) and central city quarters by P. m. brongniardii/merremius (Eastern France lineage, normal type). In the Neckar valley there is a series of mixed populations of these haplotypes and in addition of P. m. brongniardii/merremius (Eastern France lineage, Languedoc type), P. m. maculiventris (Eastern or synonymously Venetian lineage) and of P. m. nigriventris (Tuscany lineage). We present the history of introduction and the dynamics of secondary expansion based on the information available to us. Where respective data is missing we offer and discuss hypotheses on these issues.
Key words: Reptilia, Lacertidae, Podarcis muralis, introduced species, origin, urban habitats, mitochondrial DNA, Stuttgart, Germany.
Zusammenfassung
Im Stadtgebiet Stuttgart fällt eine große phänotypische Heterogenität von Mauereidechsen (Podarcis muralis) auf, die den Schluss nahe legt, dass verschiedene Standorte von allochthonen Mauereidechsen mit unterschiedlicher Herkunft besiedelt werden. Daher haben wir in den Jahren 2007 bis 2010 wichtige Standorte der Mauereidechse im Stadtgebiet untersucht. Das Ziel unserer Untersuchungen war es, die Verbreitung verschiedener Phänotypen genauer zu untersuchen und bisher fehlende Daten zur genetischen Herkunft der Mauereidechsen zu gewinnen. Wir entnahmen von zwanzig Tieren Gewebeproben und bestimmten deren Haplotyp mittels mtDNA-Analyse. Im Westen des Stadtgebiets siedeln P. m. maculiventris (westliche oder synonym Southern Alps-Linie) im Zentralbereich dagegen P. m. brongniardii/merremius (Eastern France-Linie, Normaltyp). Im Neckartal existiert eine Reihe von Mischpopulationen dieser Formen mit zusätzlich P. m. brongniardii/merremius (Eastern France-Linie, Languedoc-Typ), P. m. maculiventris (östliche oder synonym Venetian-Linie) und von P. m. nigriventris (Tuscany-Linie). Wir beschreiben die Besiedlungsgeschichte und die sekundäre Ausbreitungsdynamik von Podarcis muralis innerhalb Stuttgarts. Bei fehlenden Daten formulieren und diskutieren wir Hypothesen.
SchlГјsselbegriffe: Reptilia, Lacertidae, Podarcis muralis, allochthone Vorkommen, Herkunft, urbane Habitate, mitochondriale DNA, Stuttgart.
Einleitung
Historische Befunde
Stuttgart liegt außerhalb des natürlichen Verbreitungsgebiets der Mauereidechse. Die Vorkommen dort gehen auf Aussetzungen oder Verschleppungen zurück. Die natürliche Verbreitung der Mauereidechse in Baden-Württemberg erstreckt sich entlang des Rheins und der Unter- und Mittelläufe seiner Nebenflüsse, wobei im Gebiet des mittleren Neckars der Hohenasperg bei Ludwigsburg und Marbach am Neckar südliche Randpunkte darstellen (LAUFER et al. 2007). Die Entfernungen zum nächstgelegenen Stuttgarter Vorkommen bei Münster betragen jeweils ca. 15 km.
KUBACH & MATTHÄUS (1997) präsentieren eine Übersicht über Vorkommen von Mauereidechsen in Stuttgart. Sie beziehen sich dabei auf KLUNZINGER (1902, zitiert in BAUER 1987) und erwähnen die Aussetzung von zwölf Mauereidechsen aus dem ca. 30 km südwestlich gelegenen Wildberg a. d. Nagold in Stuttgart »in den Kriegsbergen « im Jahr 1874 durch Prof. JÄGER. Auch QUETZ (2002) und LAUFER et al. (2007) nehmen darauf Bezug. Zum Zeitpunkt der Aussetzung gab es dort noch viele Weinberge. Seit ungefähr 1900 ist das Kriegsberggebiet mit Villen und großzügigen Gärten bebaut. Lediglich ein Weinberg, der einen Blickfang im Stadtbild darstellt, ist verblieben (s. Abb. 5.). In Wildberg a. d. Nagold besteht noch heute ein individuenstarkes Vorkommen autochthoner Mauereidechsen (K. FRITZ u. P. OEFINGER schriftl.).
QUETZ (2002) realisierte eine Kartierung der Mauereidechsenvorkommen im gesamten Stadtgebiet. Die Stadt Stuttgart hat im Rahmen von Bebauungsplanänderungen mehrere unveröffentlichte ökologische Gutachten in Auftrag gegeben, in denen unter anderem auch Mauereidechsenvorkommen berücksichtigt wurden. Wir konnten darin Einsicht nehmen soweit sie die Areale am ehemaligen Güterbahnhof West (Geißeichstraße), am Weinberg an der Kielmeyer-/Fraunhoferstraße, am Travertinpark, in Wangen (Am Großmarkt) sowie den ehemaligen Güterbahnhof Bad Cannstatt (Mercedesstr./Benzstr.) und den Abstellbahnhof Untertürkheim betreffen.
Allerdings vernachlässigen alle genannten Arbeiten die Unterarten im Stadtgebiet und geben keine Auskunft zu genetischen Untersuchungen, die offenbar vor unserer Arbeit noch nicht durchgeführt wurden. Wenn früher auf die Herkunft der Tiere Bezug genommen wurde, ist hypothetisch angenommen worden, dass es sich um Nachkommen der 1874 ausgesetzten quasi-autochthonen Tiere handelte. Diese Vermutung kann aber bereits augenscheinlich widerlegt werden, da zwischen und auch innerhalb einzelner Stuttgarter Standorte die phänotypische Variabilität weit über die Variationsbreite der in Baden-Württemberg heimischen Unterart der Mauereidechse hinausgeht. Es war daher naheliegend, das phänotypisch erkennbare Spektrum durch genetische Untersuchungen zu überprüfen und abzusichern.
Ausgesetzte Mauereidechsen in Stuttgart
Nomenklatorische und genetische Anmerkungen
Die in Baden-Württemberg heimische Unterart der Mauereidechse (P. m. brongniardii Daudin, 1802) erscheint bei MERTENS & WERMUTH 1960 als brogniardii ( = error typographicus). GRUSCHWITZ & BOEHME (1986) und andere Autoren, z. B. SCHULTE (2008), übernehmen diesen Fehler, verwenden allerdings die doppelt fehlerhafte Schreibweise brogniardi. Der grammatikalisch falsche Genitiv »brongniardi« von »Brongniardius«, des latinisierten Namenspatrons Brongniart (DAUDIN hat offenbar bei der Latinisierung dieses Namens das im Französischen stumme End-T in das weichere D transskribiert), gilt jedoch nach den Internationalen Regeln der Zoologischen Nomenklatur nicht als Fehler. Dort wird lediglich empfohlen, aber nicht zwingend gefordert, die Schreibweise des erstbeschreibenden Autors zu übernehmen. Wir halten uns an diese Empfehlung.
Das natürliche Verbreitungsgebiet von P. m. brongniardii reicht von Zentral- und Ostfrankreich von der Rhone-Mündung bis in die Niederlande (Maastricht) und in die Umgebung von Aachen. Diese Unterart und P. m. merremius, welche in der Westschweiz und Westdeutschland entlang des Rheins mit Nebenflüssen bis Bonn vorkommt, haben sich nach mtDNA-Analysen als genetisch homogen erwiesen. Sie werden bis zu einer nomenklatorischen Festlegung in Anlehnung an SCHULTE et al. (2008) in dieser Arbeit als brongniardii/merremius bezeichnet da sie vom Haplotyp Eastern France (Normaltyp, d. h. der Typ mit der weitesten Verbreitung) sind. Sehr wahrscheinlich wird in Zukunft brongniardii der valide gemeinsame Unterartname sein. Innerhalb dieser Unterart wurde von der Eastern France-Linie neben dem Normaltyp noch ein SW-französischer Haplotyp Languedoc aus der Umgebung von Montpellier vabgetrennt (W. MAYER schriftl.). Der in Baden-Württemberg heimische Haplotyp ist der Normaltyp.
Um einem verbreiteten Irrtum vorzubeugen möchten wir mit W. MAYER (schriftl.) klarstellen: Unter Haplotyp ist als genetischer Typ derjenige der mitochondrialen DNA zu verstehen. Genetische Unterschiede zwischen Haplotypen können nicht für subspezifische Taxonomie eingesetzt werden. Eine Subspezies (auch Spezies) wird durch ihre Kern-DNA charakterisiert. Diese ist u. a. für die Morphologie verantwortlich. Bei Kontakt unterschiedlicher Haplogruppen kommt es sehr oft zur Introgression mitochondrialer DNA in ganz andere Morphotypen. Als Beispiel sei der Typ P. m. nigriventris-II (s. u.) aus der Umgebung von Bologna erwähnt, der morphologisch dieser Unterart entspricht, jedoch mit den Mitochondrien des P. m maculiventris-Ost Haplotyps ausgestattet ist. SCHULTE et al. (2008, 2011) trennen die Unterart maculiventris in einen West- (syn. Southern Alps) und Ost- (syn. Venetian) Haplotypen auf. Der West-Haplotyp ist im westlichen Oberitalien, der Südschweiz, Südtirol und Tirol mit einem kleinen Brückenkopf am Inn in Bayern beheimatet. Dieser Westtyp ist zusammen mit dem Haplotypen des autochthonen brongniardii/merremius Normaltyps der häufigste in allochthonen deutschen Mauereidechsenbeständen (SCHULTE et al. 2011). Der maculiventris-Osttyp ist im östlichen Oberitalien sowie an der Slowenischen und Kroatischen Küste beheimatet. Die Ost- und West-Haplotypen können auch phänotypisch unterschieden werden.
Von der Unterart P. m. nigriventris wird in SCHULTE et al. (2008, 2011) ein Typ I (Tuscany-Linie aus der Toskana, der einen kombinierten Haplo- und Phänotypen darstellt) und ein Typ II vom Nordhang des Apennins bis in die Umgebung Bolognas unterschieden. Phänotypisch sind beide Typen einander sehr ähnlich und fallen durch eine grüne Rückenfärbung und eine kontrastreiche Zeichnung auf. In einer Übergangszone vom Nordhang des Apennin bis über Bologna hinaus finden sich neben den üblichen braunrückigen P. m. maculiventris-Ost auch die oben erwähnten grünrückigen Tiere vom P. m. nigriventris Typ II. Diese grünen Tiere sind vom identischen Haplotyp wie P. m. maculiventris-Ost, weisen jedoch morphologische Charakteristika der Unterart P. m. nigriventris, also der südlich des Apennins verbreiteten Tuscany-Linie auf. Seine Entstehung erklärt W. MAYER (mündl.) durch die Hypothese einer natürlichen Introgression von männlichen Typ-I Tieren aus der Toskana über den Apennin nach der letzten Eiszeit und einer Hybridisierung mit weiblichen P. m. maculiventris-Ost-Tieren. Bekannte Vorkommen nördlich der Alpen von P. m. nigriventris II sind die allochthonen Populationen um Passau, Dresden und Mannheim (SCHULTE et al. 2011). Die Tiere an den genannten Orten wurden früher wie die Tiere des Typs I aus der Toskana als Podarcis muralis brueggemanni bezeichnet (z. B. in MERTENS & WERMUTH 1960). Die Terra typica von P. m. brueggemanni (La Spezia) liegt außerhalb der Toskana in Ligurien. Die Tiere von dort bilden einen eigenen kombinierten Haplo- und Phänotyp (W. MAYER schriftl.). Da wir diesen Typ in Stuttgart nicht nachweisen konnten, gehen wir nicht weiter darauf ein.
Wir haben es also mit der verwirrenden Situation zu tun, dass sich einerseits zwei bisher valide Unterarten (brongniardii und merremius) in einer genetischen Linie (Eastern France) wiederfinden, diese sich wiederum in zwei Subtypen (Normal und Languedoc) aufspaltet, welche aber nicht den bisher validen Unterartnamen entsprechen. Andererseits wurde eine bisher valide Unterart (maculiventris) in zwei deutlich getrennte Haplo- und Phänotypen (West, syn. Southern Alps und Ost, syn. Venetian) aufgespalten. Zusätzlich verkompliziert sich die Sachlage dadurch, dass die natürliche Einkreuzung von grünen nigriventris I-Männchen in maculiventris-Ost-Bestände nur phänotypische Spuren – unter anderem die Grünfärbung – hinterlässt. Der mtDNAHaplotyp, der nur die mütterliche Linie des Erbgangs widerspiegelt, bleibt aber unbeeinflusst. Dieser umfänglich erscheinende Exkurs über Haplo- und Phänotypen erscheint uns zum Verständnis des Unterartspektrums und der genetischen Linien der Stuttgarter Mauereidechsen hilfreich, namentlich für Feldherpetologen, die mit Fragen der molekularen Systematik nicht vertraut sind.
Methoden
Neben Literaturangaben dienten vor allem Hinweise von Einzelpersonen als Informationsquellen. Schon bekannte und durch Auskünfte neu hinzugewonnene Stuttgarter Standorte von Mauereidechsen wurden in der Zeit vom Frühjahr 2007 bis Herbst 2010 bei geeigneter Witterung mehrfach aufgesucht. Dabei wurden von Mauereidechsen bevorzugte Habitate gezielt begangen. Gesichtete Eidechsen (Mauereidechsen und Zauneidechsen) wurden registriert und fotografisch dokumentiert. Lokale Populationsgrößen wurden in Anlehnung an die Laufersche Faustregel (LAUFER 1998) »Anzahl gesichteter Adulttiere bei zweimaligem Abgehen eines Transsekts mal vier und proportionale Hochrechnung auf die nach Augenschein besiedlungsfähigen Teile des Gesamthabitats« grob geschätzt. Diese Schätzung liefert eine untere Schranke für die Gesamtzahl von Individuen eines Habitats; dies haben unsere Beobachtungen in Wangen (s.u.) bestätigt. In die Schätzungen der Gesamtzahl adulter Tiere in den drei Siedlungsbereichen West, Zentral und Neckartal (s. u.) flossen neben eigenen Beobachtungen auch Literaturangaben und Hinweise von Einzelpersonen ein.
Zur genetischen Untersuchung der Mauereidechsen wurden entsprechend zweier Ausnahmegenehmigungen des Regierungspräsidiums Stuttgart nach §43 Abs. 8 Ziff. 3 BNatSchG insgesamt 20 Tiere beprobt. Hierfür wurden einzelne Mauereidechsen mit der Hand oder einer Schlinge gefangen. Das Schwanzende gefangener Tiere diente als Probenmaterial und wurde ungefähr 1 cm vor der Spitze an einer Sollbruchstelle vorsichtig abgedreht, um eine nachfolgende Regeneration zu ermöglichen. Bei chirurgischer Durchtrennung eines Wirbelkörpers verheilt die Wunde nur und es wird kein Regenerat ausgebildet (C. BENDER mündl.). Nach der Beprobung wurden die Tiere sofort wieder am Fundort freigelassen. Die Schwanzspitzen wurden in Eppendorf-Küvetten mit 70 %igem Ethanol fixiert und bis zum Versand bei Raumtemperatur gelagert.
Das Analyseverfahren in den Untersuchungslaboratorien (WERNER MAYER, 1. Zoologische Abteilung des Naturhistorischen Museums Wien (18 Proben) und Abteilung Biogeografie der Universität Trier (3 Proben, davon 1 Doppelbestimmung)) ist bei SCHULTE et al. (2008) beschrieben. Da der Haplotyp mittels eines Abschnitts (650–887 bp) des mitochondrialen Cytochrom-b-Gens bestimmt wurde, kann nur die Herkunft der mütterlichen Linie des jeweiligen Individuums nachgewiesen werden.
Zuordnungen erfolgten durch WERNER MAYER nach dem umfangreichen Wiener Datensatz aus einer bislang unpublizierten Studie (SCHWEIGER et al. unveröff.).
Ergebnisse
Die Mauereidechse besiedelt in Stuttgart erwartungsgemäß Sekundärhabitate wie Weinberge, Steinbrüche, betriebene oder stillgelegte Bahnanlagen, Ruderalgelände sowie steile Strukturen wie Mauern oder Hangbereiche mit Staffeln (lokale Bezeichnung für Außentreppen) und dekorativen Steinstrukturen in Hausgärten und gärtnerisch gestalteten Anlagen. In Stuttgart können drei größere von Mauereidechsen besiedelte Bereiche definiert werden. Dazu haben wir auf der Grundlage unserer genetisch gesicherten punktuellen Befunde die Bereichsgrenzen durch Extrapolation in Übereinstimmung mit Sichtbeobachtungen von Tieren und deren Phänotyp sowie der Eignung des Geländes als Mauereidechsenhabitat nach Augenschein eingeschätzt.
a) Der westliche Siedlungsbereich mit Beprobungsorten W1–W3 (Abb. 1) erstreckt sich in einem bis zu 2000 m breiten Streifen beidseits der Trasse der Gäubahn südwestlich des Westportals des Kriegsbergtunnels. Dieses Areal umfasst ca. 4,5 km2 und beherbergt nach unserer Schätzung mehr als 1000 adulte Mauereidechsen. Unmittelbar entlang der Gleisbereiche besteht ein nahezu geschlossenes Vorkommen und auf dem Birkenkopf (W3) ein Isolat in einem von Straßen durchschnittenen Waldgebiet.
 |
|
|
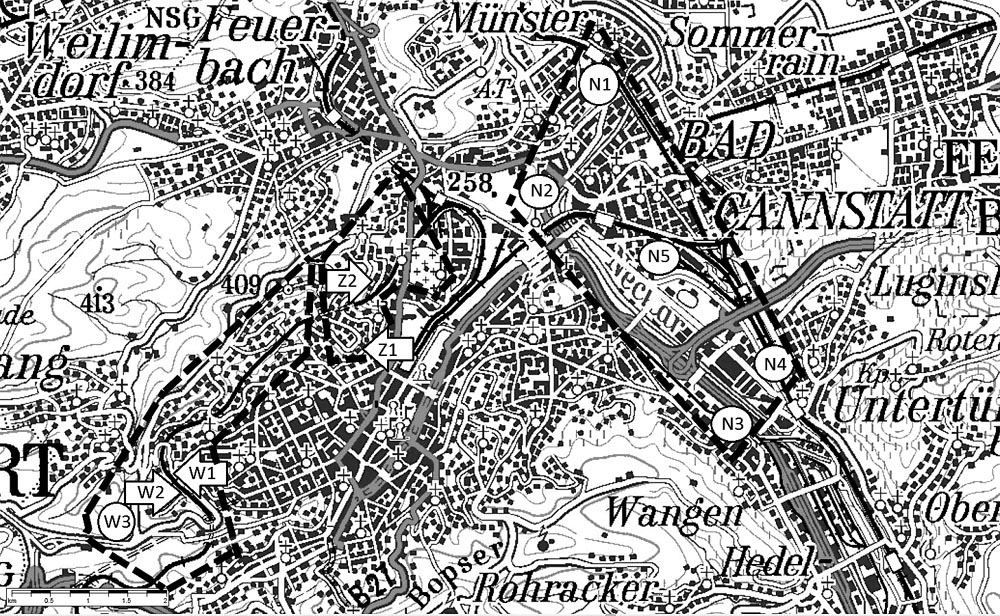
Abb. 1: Abb 1: Geschätzte Grenzen der Bereiche West (Beprobungsorte W1–W3), Zentral (Z1–Z2) und Neckartal (N1–N5). Ellipsen: Mehrfachbeprobungen, Pfeile: Einzelbeprobungen. Die Ortsabkürzungen werden im Text näher erläutert und in Tabelle 1 wird darauf Bezug genommen. Maßstabslineal: 2 km. Quelle: Digitale topografische Karte 1 : 200000 des Landesvermessungsamtes Baden-Württemberg, 2006.
Estimated boundaries of the areas West (sampling locations W1–W3), Central (Z1–Z2) and Neckar Valley (N1–N5). Ellipses: multiple samplings, arrows: single samplings. Abbreviations of locations are explained in the text and referred to in fig 1. Scale rule: 2 km. Source: Digital topographic map 1 in 200,000, edited by the Board of Land Survey of the federal state of Baden-Württemberg, 2006.
|
Die übrigen Standorte liegen vereinzelt meist in privaten Hausgärten oder an öffentlichen Staffeln durch gärtnerisch gestaltete Hangbereiche. Einwanderungen nach Botnang und Feuerbach können wir nicht ausschließen; jedoch wurden diese Stadteile von uns nicht untersucht. Der Haplotyp P. m. maculiventris-West wurde entlang eines Querschnittstranssekts anhand je eines Tieres vom Herderplatz (Ort W1, Abb. 1, 2) und von der Eisenbahnbrache des ehemaligen Güterbahnhofs West (Ort W2, Probe GBW1) sowie von zwei Tieren vom Birkenkopf (Ort W3, Abb. 1, 3, 4) bestimmt. Phänotypisch ergaben sich im westlichen Bereich keine Hinweise auf andere Unterarten oder genetische Linien.
 |
|
|
Abb. 2: Staffel zur U-Haltestelle Herderplatz (W1). Foto: G. DEICHSEL. Stair to subway stop Herderplatz.
|
 |
|
|
Abb. 3 (links): Birkenkopf (W3) Untere sГјdexponierte Windung der FahrstraГџe. Abb. 4 (rechts): Birkenkopf (W3) Mittlere sГјdexponierte Windung der FahrstraГџe. Fotos: G. DEICHSEL.
Left: Birkenkopf hill (W3) Lower south-exposed winding of driveway. Right: Birkenkopf hill (W3) Middle south-exposed winding of driveway.
|
b) Der zentrale Siedlungsbereich erstreckt sich vom Kriegsberggebiet (Ort Z1, Abb. 1, 5) über die Weinberge an der Mönchhalde und an der Kielmeyer-/ Fraunhoferstraße (Ort Z2, Abb. 1, 6) bis ungefähr zum Pragsattel im Norden der Kernstadt. Er umfasst ca. 2,1 km² und wird von P. m. brongniardii/merremius (Normaltyp) besiedelt. Davon abweichende Phänotypen konnten wir nicht finden. Einwanderungen nach dem Stadtteil Feuerbach können wir nicht ausschließen. Möglicherweise reicht der Zentralbereich auch bis in die Nähe des Travertinparks im Stadtteil Bad Cannstatt an der Grenze zum Stadtteil Münster (Ort N1, Abb. 7), da wir dort unter anderem auch denselben Haplotypen nachweisen konnten. Die Verbreitung im Zentralbereich ist lückenhaft und außerhalb der gehäuften Vorkommen in und um Weinberge schwierig zu erfassen, da sehr oft unzugängliche private Hausgärten besiedelt werden. Wir schätzen den Bestand im Zentralbereich auf mehr als 500 Tiere. Die Verläufe möglicher Kontaktzonen zur Population von P. m. maculiventris-West um die Gäubahntrasse am Westportal des Kriegsbergtunnels und zu der Mischpopulation im Travertinpark konnten wir im Rahmen der hier beschriebenen Untersuchung nicht erfassen.
 |
|
|
Abb. 5: Kriegsberggebiet (Z1) vom Bahnhofsturm aus. Historische Aufnahme von 2008 vor Beginn der Abbrucharbeiten zur Umgestaltung des Stuttgarter Hauptbahnhofs. Foto: G. DEICHSEL.
Kriegsberg district (Z1) as viewed from the Central Railway Station tower. Historic photograph of the year 2008 before the demolition works for the re-construction of the station building.
|
 |
|
|
Abb. 6 (links): Weinberg (Z2) an der Kielmeyer-/FraunhoferstraГџe. Abb. 7 (rechts): Ehemaliger Steinbruch im Travertinpark (N1). Fotos: G. DEICHSEL.
Left: Vineyard at Kielmeyer/Fraunhofer street. Right: Former quarry in Travertin Park (N1).
|
QUETZ (2002) verzeichnet einen Standort am Pragsattel in der möglichen Kontaktzone zum Travertinpark, den wir bei einer einmaligen Begehung nicht bestätigen konnten. Im Rosensteinpark, der eine potenzielle Kontaktzone zu der Mischpopulation in und an der Wilhelma (Ort N2, Abb. 8) darstellen könnte, wiesen wir keine Mauereidechsen nach.
 |
|
|
Abb. 8: Parkanlage an der Wilhelma-Schifflände (N2) mit GD (l) und AK (r). Die geschwungenen Sitzbänke werden von den Mauereidechsen als Sonnenplätze genutzt. Foto: A. CONSUL.
Park at Wilhelma boat stop with GD (l) and AK (r). The curved benches serve as basking perches for the wall lizards.
|
|



